

 当前位置:首页 > 专题 > 分子生物学
当前位置:首页 > 专题 > 分子生物学
病毒载体是基因体外实验与动物活体造模常用工具,主流包含慢病毒、腺病毒、腺相关病毒(AAV)、逆转录病毒四类,本文分别从病毒结构、基因组、侵染原理、产品特点逐一介绍,附带四大载体参数对比表格,方便科研选型。
慢病毒是一种有包膜的RNA病毒,直径为80-120nm,呈二十面体对称结构、球形。病毒颗粒最外层是包膜(包膜蛋白决定了感染细胞的类型),再往里依次为基质蛋白和衣壳,最里面是两条相同的正股RNA链和酶(逆转录酶、整合酶和蛋白酶)。
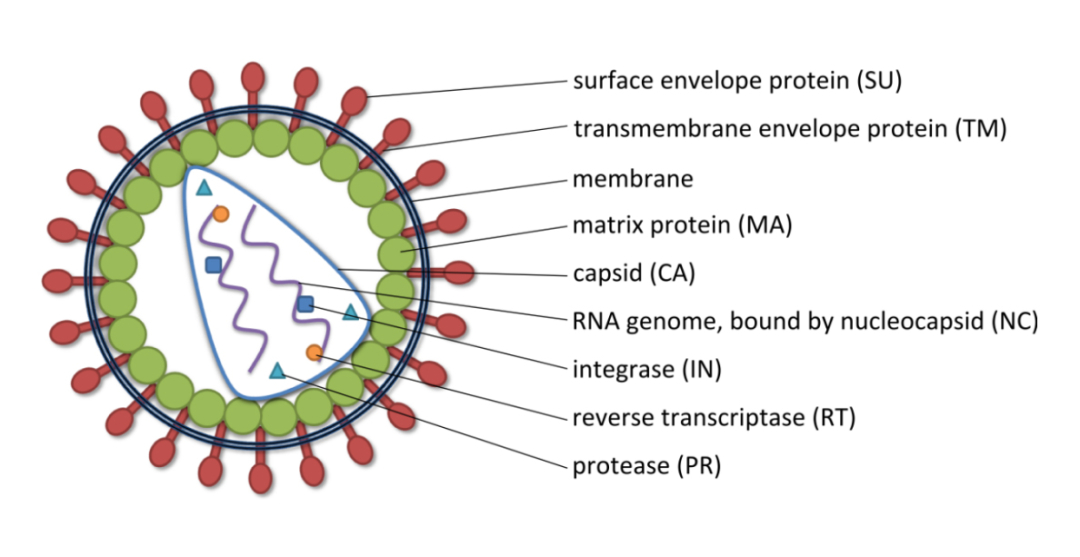
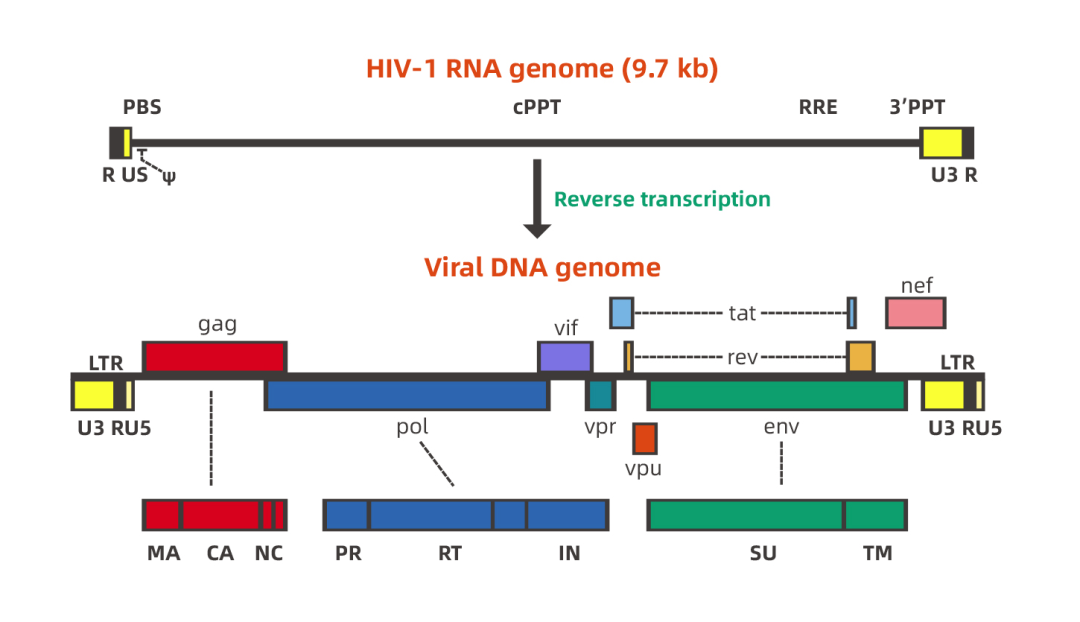
慢病毒含有复杂的基因组。以HIV-1(人类免疫缺陷I型病毒)为例,HIV是一种单链RNA病毒,其基因组长度约为9.7kb。基因组两端各有一个反向末端重复序列(inverted terminal repeat, ITR),中间包括gag、 pol、env 3个结构基因及tat、rev、nef、vif、vpr、vpu 6 个调节基因。
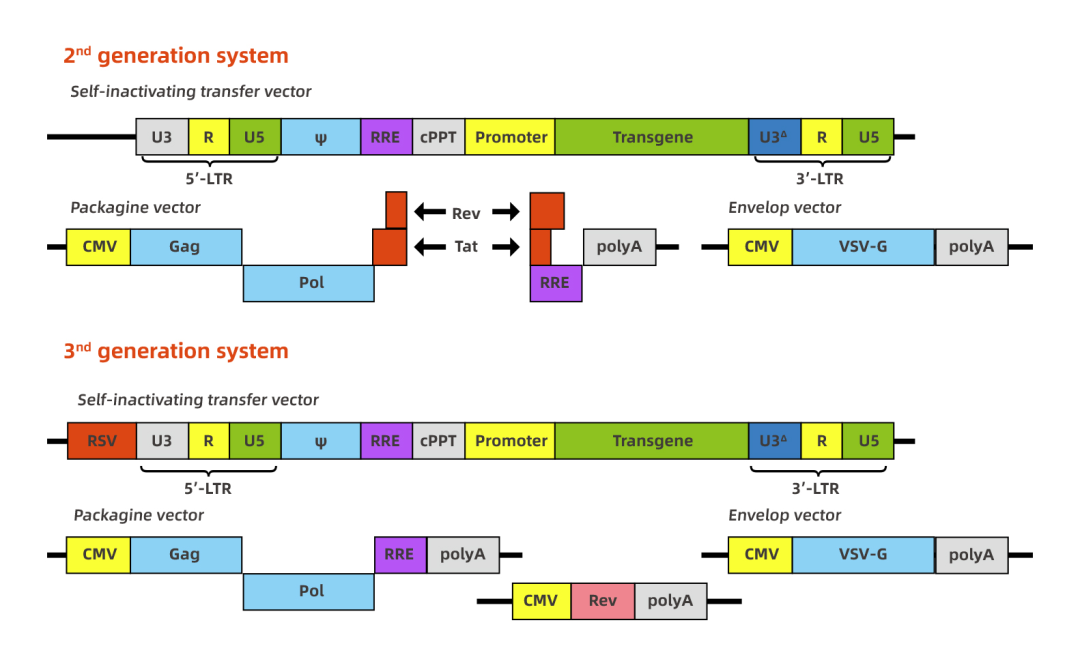
慢病毒是以HIV-1为基础发展起来的基因治疗载体。慢病毒载体的研究发展得很快,研究的也非常深入。出于包装能力和安全性考虑,目前慢病毒载体已发展了很多代,其中2代和3代是目前大家应用比较广的。慢病毒载体的包装容量为7-8kb,但是通常超过3kb的话会明显影响慢病毒的滴度。
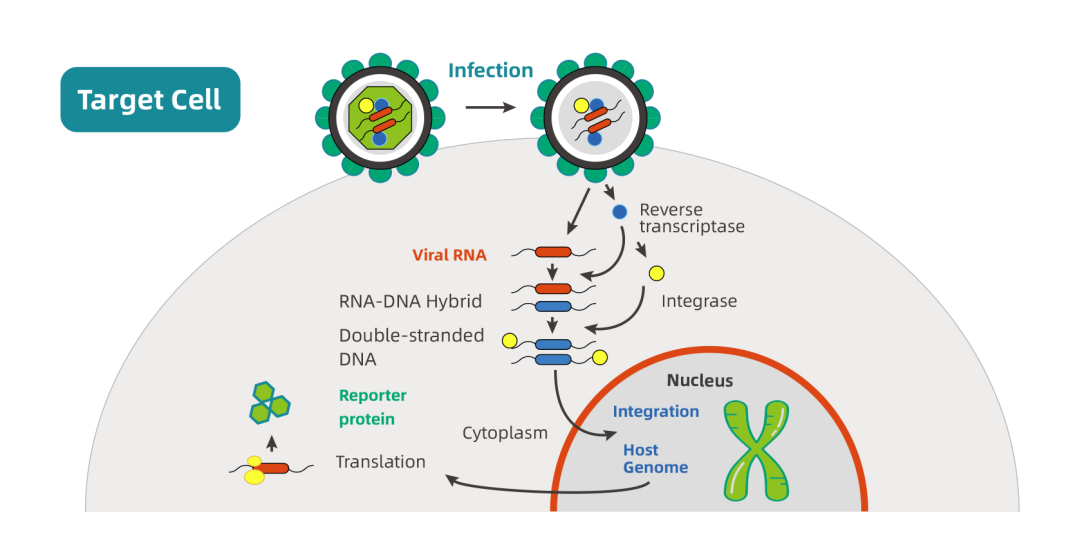
慢病毒感染细胞的第一步是通过包膜蛋白与细胞表面受体结合。慢病毒与宿主细胞膜融合后释放结构蛋白、酶蛋白和病毒核心。病毒RNA在逆转录酶的作用下逆转录并与整合酶形成整合前复合物。整合前复合物进入细胞核后,整合酶催化其整合至宿主基因组。外源基因前启动子驱动其在细胞质中表达。
1. 慢病毒有广泛的宿主范围,能够有效地感染分裂细胞、非分裂细胞,特别适合于一些难转染的细胞(如原代细胞、干细胞、不分化的细胞)和对腺病毒感染具有较强免疫反应的细胞(如树突状细胞、单核细胞和间充质干细胞)等,能大大提高目的基因转导效率,而且目的基因整合到宿主细胞基因组的几率大大增加。
2. 慢病毒能将外源基因有效地整合入宿主染色体上,介导目的基因稳定、长期的表达。因此,可以利用慢病毒建立稳定细胞株。如需定制载体可选用慢病毒包装服务。
3. 适用范围更广,不仅能用于体外实验,还能用于活体动物模型,尤其是动物成瘤实验。
腺病毒是一种直径为90-100nm的病毒颗粒,衣壳呈廿面体,由240个六邻体和12个五邻体组成。以五邻体蛋白为基底由衣壳表面伸出12根纤毛,纤毛顶端形成头节区。五邻体和纤毛的头节区可与细胞表面的受体结合,在病毒感染细胞过程中起着非常重要的作用。
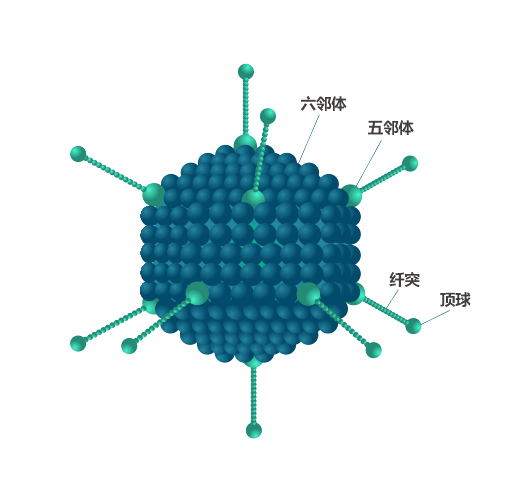
腺病毒是一种双链线性的DNA病毒,其基因组长度约为36kb。基因组两端各有一个103 bp的反向末端重复序列(inverted terminal repeat, ITR),参与病毒DNA的复制;ITR的内侧为病毒包装信号Ψ,参与腺病毒基因组的衣壳化。基因组包含早期表达的与腺病毒复制相关的E1~E4基因和晚期表达的与腺病毒颗粒组装相关的L1~L5基因。
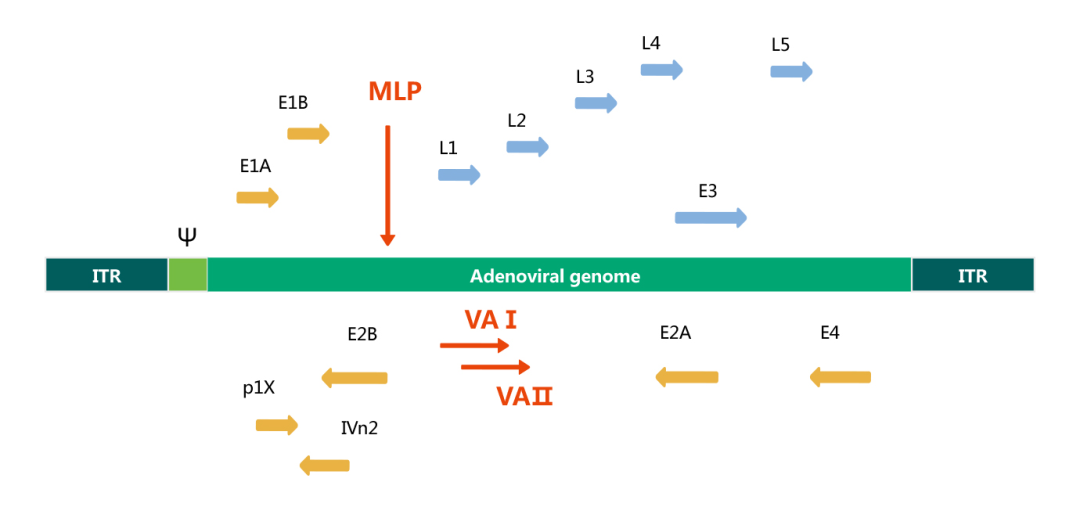
基于人血清5型腺病毒的基因组结构,结合各基因的功能,科学家们开发了缺失E1和E3基因的Ad5腺病毒载体。E1基因在组装感染性病毒颗粒时必不可少,但是可以在HEK293包装细胞中得到补充,而E3基因不影响病毒的包装。由于E1和E3基因的缺失,腺病毒载体可插入高达7.5kb的外源基因。
腺病毒感染细胞的第一步是通过纤突蛋白与细胞表面CAR结合。腺病毒粘附后,五邻体蛋白上的精氨酸-谷氨酸-天冬氨酸(RGD)与细胞表面的整合素αvβ3和αvβ5结合并相互作用使腺病毒进入细胞。
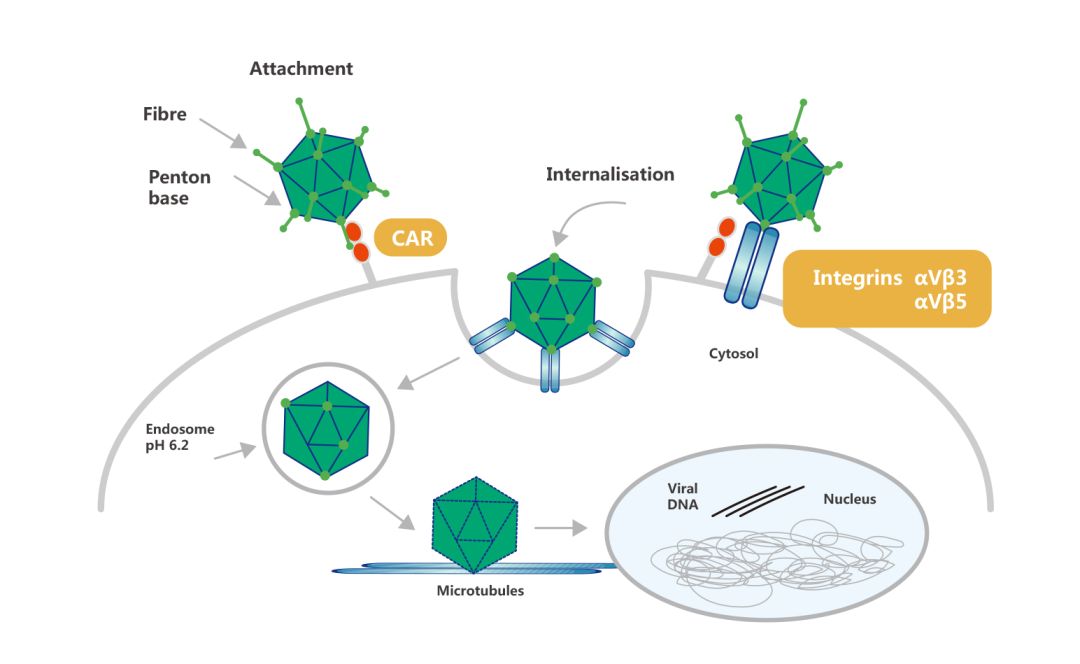
1.腺病毒感染的宿主细胞范围广,包括分裂细胞和不分裂细胞(某些抗腺病毒感染的淋巴瘤细胞除外)。
2.腺病毒感染效率高,在最佳感染条件下,感染率可达100%。
3.腺病毒的病毒滴度可以很高,纯化、浓缩后可达10¹²⁻¹³vp/ml(即10¹⁰⁻¹¹pfu/ml)。
4.腺病毒易于扩增,而其他病毒比如慢病毒和腺相关病毒需要重新包装。
5.不整合到染色体中,无插入致突变性。
6.腺病毒理化性质稳定,4℃:数周;-80 ℃:数年。科研构建可选择腺病毒载体包装服务。
腺相关病毒(Adeno-associated virus)属于细小病毒科,Dependoparvovirus属,是目前发现的一类结构简单、无包膜的单链DNA缺陷型病毒,它的生命周期依赖于复制病毒的参与,如腺病毒和单纯疱疹病毒。直径在20-26nm范围内,其直径为腺病毒的1/5,慢病毒的1/4,呈现20面体结构,对分裂细胞和非分裂细胞均可感染。
腺相关病毒含有约4.7kb的单链线性DNA基因组。基因组两端为末端反向重复序列(ITR),对于病毒的复制和包装具有重要作用;中间基因组编码两类蛋白Rep和Cap,Rep蛋白参与病毒的复制和整合,Cap蛋白为病毒衣壳蛋白。衣壳包含三种类型的亚基VP1、VP2和VP3,比例为1:1:10(VP1:VP2:VP3)。基于AAV的基因组结构,基因工程化的AAV载体删除了全部AAV蛋白编码序列,添加了外源基因的表达盒。唯一的病毒来源序列是ITR,它们是在病毒包装过程中指导AAV复制和包装所必需的。病毒编码序列的完全去除使AAV的包装能力最大化,并且有助于它们在体内递送时的低免疫原性和细胞毒性。
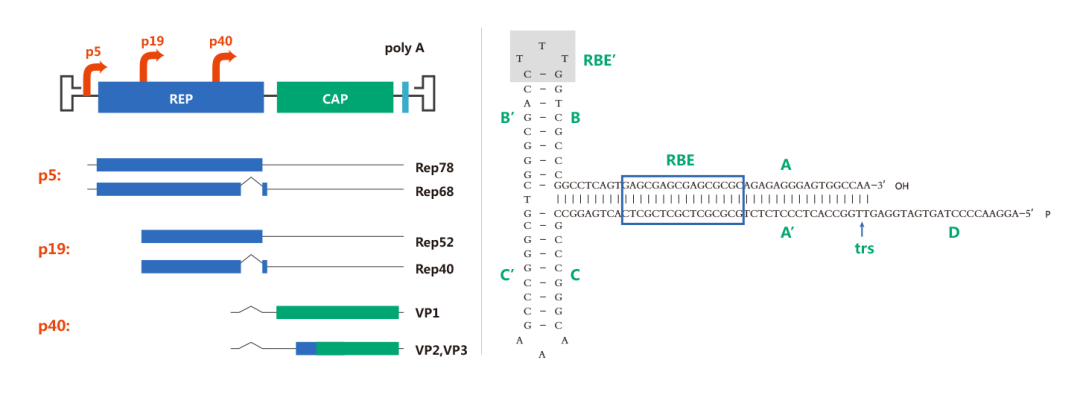
现阶段研究人员已发现12种人类AAV 血清型(AAV1至AAV12)和 100多种非人类灵长动物AAV血清型。不同AAV血清型具有不同的衣壳蛋白空间结构、序列和组织特异性,因而其识别与结合的细胞表面受体也相应有很大差别,这也导致不同血清型转染的组织类型、细胞类型和感染效率也各不相同。
下表汇总AAV1~AAV9血清型受体及组织嗜性,是AAV载体选型重要参考。
表1 人类AAV1至AAV9识别的细胞表面受体
| Serotype | Glycan recognitiona | Coreceptor |
|---|---|---|
| AAV1 | Neu5Aca2-3GalNAcβ1-4GlcNAc | Unknown |
| AAV2 | 6-O-and N-sulfated heparin | Fibroblast / hepatocyte growth factor receptor;laminin receptor; integrin αVβ5 and α5β1 |
| AAV3 | 2-O-and N-sulfated heparin | Hepatocyte growth factor receptor; Laminin receptor |
| AAV4 | Galβ1-4GlcNAcβ1-2Manα1- 6Manβ1-4GlcNAcβ1-4GlcNAc | Unknown |
| AAV5 | Neu5Acα2-3(6S)Galβ1-4GlcNAc | Platelet-derived growth factor receptor |
| AAV6 | Neu5Acα2-3GalNAcβ1-4GlcNAc; N-sulfated heparin | Epidermal growth factor receptor |
| AAV7 | Unknown | Unknown |
| AAV8 | Unknown | Laminin receptor |
| AAV9 | Galactose | Laminin receptor |
表2 AAV1至AAV9的不同组织亲噬性
| Tissue | Optimal Serotype |
|---|---|
| CNS | AAV1,AAV2,AAV4,AAV5,AAV8,AAV9 |
| Heart | AAV1,AAV8,AAV9 |
| Kidney | AAV2 |
| Liver | AAV7,AAV8,AAV9 |
| Lung | AAV4,AAV5,AAV6,AAV9 |
| Pancreas | AAV8 |
| Photoreceptor Cells | AAV2,AAV5,AAV8 |
| RPE(Retinal Pigment Epithelium) | AAV1,AAV2,AAV4,AAV5,AAV8 |
| Skeletal Muscle | AAV1,AAV6,AAV7,AAV8,AAV9 |
AAV转导细胞的过程起始于识别细胞表面糖基化修饰的受体,由网格蛋白介导的细胞内吞触发AAV内化入细胞胞浆,进入内体。AAV在细胞骨架蛋白网络的帮助下由细胞胞浆向细胞核运输。在内体的酸性环境下,AAV的衣壳蛋白构象发生变化。AAV逃离内体后或者由蛋白酶体进行蛋白降解,或者入核和脱衣壳,双链转化,进而形成环形附加体游离于细胞核。
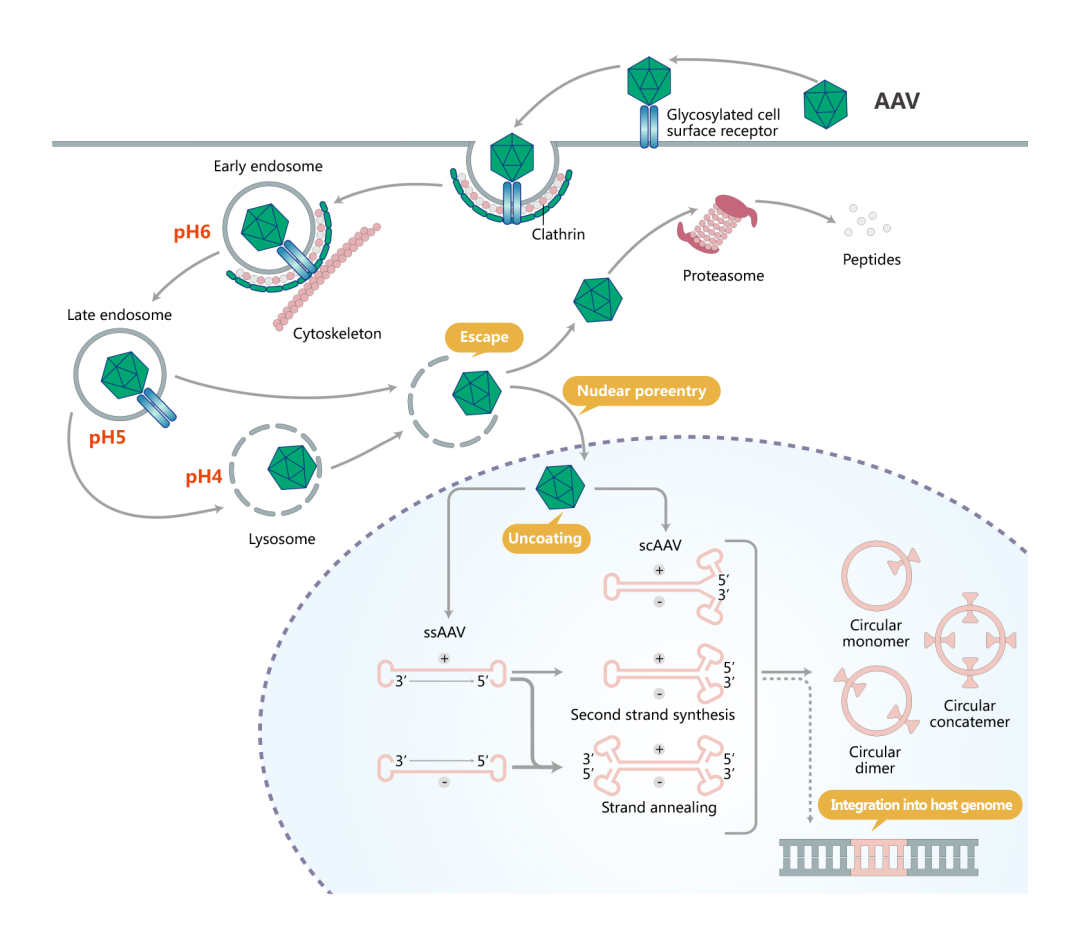
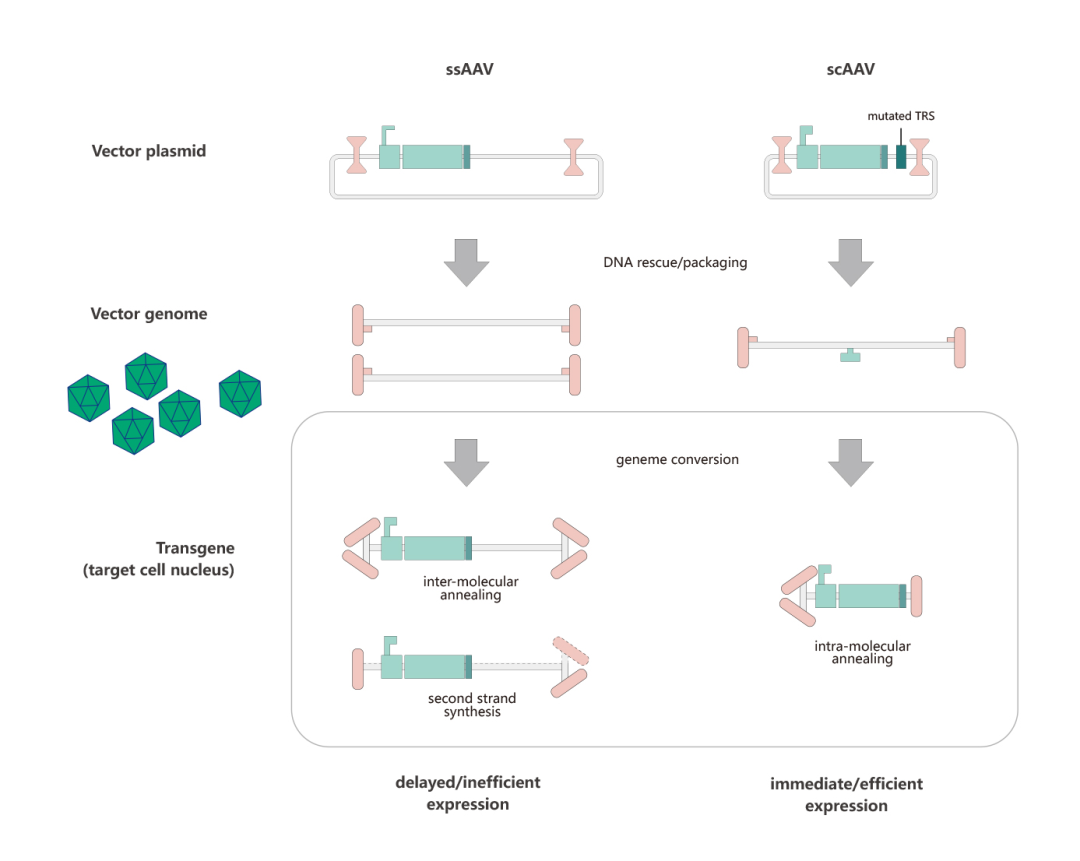
目前,研究人员常用的两类AAV分别为 single-stranded AAV (ssAAV) 和self-complementary AAV (scAAV)。scAAV是基于2个基础被开发的:一个是理论基础:无论scAAV还是rAAV,病毒颗粒中均可包裹二倍体,甚至四倍体的AAV基因组DNA;一个是结构基础:wtAAV ITR序列的特殊性(T型结构,见AAV基因组部分)。ssAAV包装基因组正义链和反义链的几率一样。ssAAV在入核、脱衣壳后,需要借助宿主DNA聚合酶或者分子间退火完成双链转化,才能启动基因转录过程,而scAAV中已存在双链,它入核后即可启动基因转录,跨过了双链转化的步骤,从而实现外源基因的快速表达。维真生物可提供AAV病毒包装、scAAV定制构建。
逆转录病毒(Retrovirus),又称反转录病毒,是一类RNA病毒,需在逆转录酶的作用下首先将RNA转变为cDNA,新合成的cDNA插入宿主的核DNA中,随宿主DNA复制、转录、翻译达到扩增目的。逆转录病毒科可分为7个属:α逆转录病毒属、β逆转录病毒属、γ逆转录病毒属、δ逆转录病毒属、ε逆转录病毒属、慢病毒属和泡沫病毒属。逆转录病毒通常指的是称为γ逆转录病毒,包括MMLV(莫洛尼氏鼠白血病逆转录病毒)和MSCV(鼠干细胞病毒)。
逆转录病毒载体通常是在γ逆转录病毒的基础上改造获得,将逆转录病毒中与病毒包装和转导相关的基因删除,替换为需要表达的外源基因,因此生产的病毒颗粒是复制缺陷型的,无法在细胞内复制并组装为新的病毒颗粒,但能将外源基因整合到宿主基因组中持续表达。逆转录病毒载体的感染过程大致包括以下步骤:
病毒感染宿主细胞:逆转录病毒载体首先通过病毒表面的包膜蛋白与宿主细胞表面的受体结合,通过胞吞进入细胞内部。
病毒RNA逆转录为DNA:进入宿主细胞后,病毒的RNA基因组被反转录酶转录为双链DNA。
病毒DNA整合到宿主基因组:逆转录病毒载体的DNA通过整合酶的作用插入宿主细胞的基因组中。
外源基因的表达:病毒DNA整合到宿主基因组后,外源基因就会受到宿主细胞内的转录和翻译机制的控制,产生所需的蛋白质或执行其他功能。
高效转导:逆转录病毒具有广泛的宿主范围,能够在分裂细胞中进行高效的基因转导。但其无法穿过核膜,只能在细胞分裂过程中通过不完整的核膜进入细胞核,并将外源基因整合到基因组中,因此只能感染分裂期的细胞。
基因整合:逆转录病毒能将外源基因插入宿主细胞基因组,因此转导后的基因能够稳定传递给子代细胞,适合用于长时间的基因表达。但病毒基因组的整合位点难以预测,可能导致潜在的副作用,如基因组不稳定或插入突变。如需载体构建可选用逆转录病毒包装服务。
综上,逆转录病毒凭借稳定整合的优势多用于构建稳转细胞株,受限于只能侵染分裂细胞,常与慢病毒、腺病毒、AAV搭配选型实验。
对于转染困难的细胞,科研工作者通常会选择病毒来导入目的基因。目前,慢病毒、腺病毒、腺相关病毒和逆转录病毒是常用的病毒载体工具。那么如何选择适合您实验体系的病毒工具,请参考下面四种病毒工具的比较:下表从粒径、基因组、包装容量、滴度多维度对比四种常用病毒载体关键参数。
| 病毒表达系统 | 腺病毒 | 腺相关病毒 | 慢病毒 | 逆转录病毒 |
|---|---|---|---|---|
| 大小 | 70-100nm | 20-26nm | 80-120nm | 80-120nm |
| 病毒外壳 | 无 | 无 | 具有包膜蛋白 | 具有包膜蛋白 |
| 基因组 | dsDNA | ssDNA | ssRNA | ssRNA |
| 基因组大小 | 38-39kb | 5kb | 9kb | 7-12kb |
| 货物基因容量 | 8kb | 5kb | 8kb | 8kb |
| 宿主种类 | 分裂/非分裂细胞 | 分裂/非分裂细胞 | 分裂/非分裂细胞 | 分裂细胞 |
| 宿主整合 | × | × | √ | √ |
| 表达丰度 | 高水平 | 高水平 | 中到高水平 | 高水平 |
| 长时间表达 | × | √ | √ | √ |
| 免疫原性 | 高 | 极低 | 中等 | 中等 |
| 相对病毒滴度 | 1010pfu/mL | 1013VG/mL | 108TU/mL | 108TU/mL |
| 生物安全等级 | BSL-2 | BSL-2 | BSL-2 | BSL-2 |
综上,根据实验细胞类型(分裂/非分裂细胞)、体内体外实验、目的基因片段大小可灵活挑选适配病毒载体,维真生物拥有成熟的慢病毒、腺病毒、AAV载体包装平台,可按需定制载体构建。

400-077-2566

service@wzbio.cn








