当前位置:首页 > 专题 > 细胞自噬
当前位置:首页 > 专题 > 细胞自噬
1.细胞自噬简介
细胞自噬(autophagy=self-eating)意为自体吞噬,是真核细胞在细胞自噬相关基因(autophagy related gene,Atg)的调控下利用溶酶体降解自身细胞质蛋白和受损细胞器的过程。细胞自噬可防止细胞损伤,促进细胞在营养缺乏的情况下存活,并对细胞毒性刺激作出反应。细胞自噬包括生理条件下的基础型细胞自噬和应激条件下的诱导型细胞自噬。前者是细胞的自我保护机制,有益于细胞的生长发育,保护细胞防止代谢应激和氧化损伤,对维持细胞内稳态以及细胞产物的合成、降解和循环再利用具有重要作用;但细胞自噬过度可能导致代谢应激、降解细胞成分,甚至引起细胞死亡等。研究表明,细胞自噬能在细胞稳态、衰老、免疫、肿瘤发生及神经退行性疾病等多种生理病理过程中发挥重要作用。
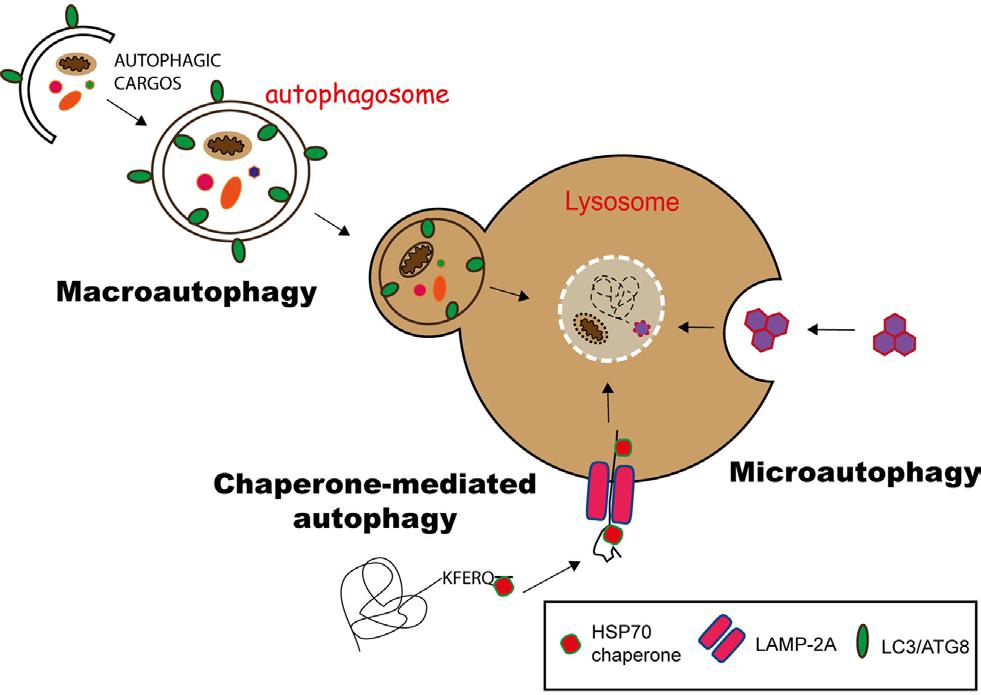
根据包裹物质及运送方式的不同可将细胞自噬分为3种类型(图1):
① 巨细胞自噬(macroautophagy):通过形成具有双层膜结构的细胞自噬体(autophagosome)包裹胞内物质,最终细胞自噬体与溶酶体融合。一般情况下所说的细胞自噬是指巨细胞自噬。
② 微细胞自噬(microautophagy):通过溶酶体或液泡表面的形变直接吞没特定的细胞器。
③ 分子伴侣介导的细胞自噬(chaperone-mediated autophagy, CMA):具有KEFRQ样基序的蛋白在HSP70伴侣的帮助下,通过LAMP-2A转运体转运到溶酶体。
|
|
图1. 3种细胞自噬途径 (Duraes Fernanda V et al. Front Immunol, 2015)
2.细胞自噬的发生过程
细胞自噬是一种在进化上保守的细胞内分解代谢过程,在该过程中,细胞质大分子、聚集性蛋白、受损细胞器或病原体被运送至溶酶体,并被溶酶体水解酶降解,产生核苷酸、氨基酸、脂肪酸、糖和三磷酸腺苷,最终再循环到胞浆中。诸如饥饿、辐射、缺氧、细菌入侵、生长因子匮乏等多种因素均可诱导细胞自噬发生。
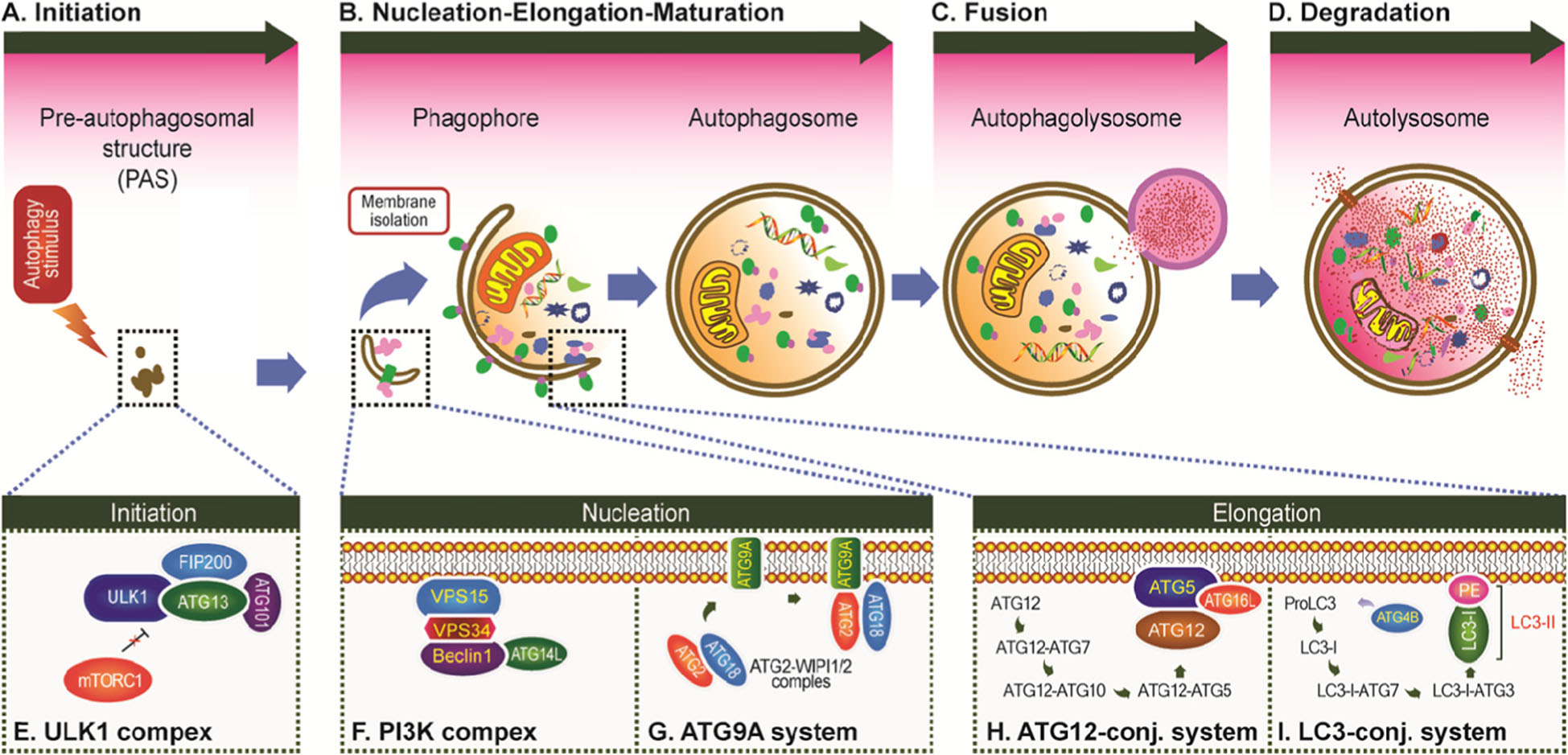
细胞自噬的发生过程大体分为以下4个阶段(图2):
①细胞自噬的起始 在细胞自噬诱导信号的调控下,ULK1复合物和多种ATG蛋白被活化,并定位于前细胞自噬体处。
②隔离膜和细胞自噬体的形成 ATG蛋白和脂质不断被募集,从而形成杯状的双层膜结构(隔离膜,phagophore);随着隔离膜的逐渐延伸,将要被降解的胞浆成分完全包裹,最终形成闭合的细胞自噬体(autophagosome)。
③细胞自噬体与溶酶体融合 细胞自噬体形成后将其包裹物通过胞内运输系统运输至溶酶体,并与溶酶体融合。
④细胞自噬体的裂解 细胞自噬体与溶酶体融合后形成细胞自噬溶酶体(autolysosome),最终在溶酶体水解酶的作用下降解其包裹物。
总而言之,细胞自噬的本质其实是细胞内的膜重排。在膜重排的过程中,会形成一个具有双层膜并裹挟着随机或特定底物的封闭囊泡,即细胞自噬体,进而与溶酶体融合,形成细胞自噬溶酶体并降解底物。
|
|
图2. 细胞自噬的发生过程 (Li et al. Molecular Cancer,2020)
3.细胞自噬相关蛋白
细胞自噬的发生与营养状态、能量状态、氧化应激、缺血缺氧等相关,其受多重机制调节,如ULK1通路、Beclin1通路、AMPK通路等。在细胞自噬的发生过程中,有多种细胞自噬相关蛋白可调节和控制细胞自噬形成的不同阶段。
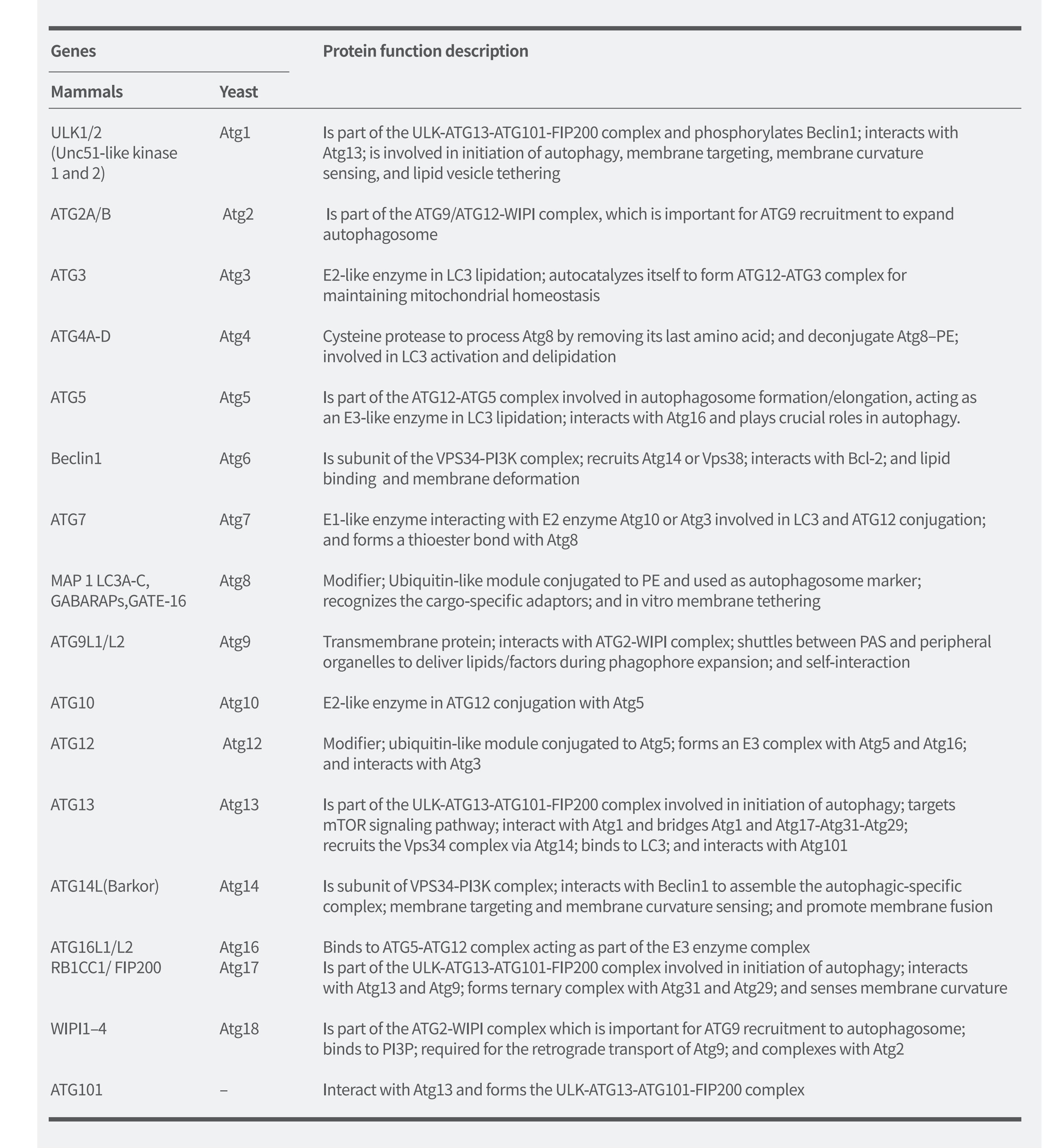
如图2所示,在细胞自噬的发生过程中,多种细胞自噬相关基因调控细胞自噬流的不同阶段。迄今为止,科学家已在酵母中鉴定出40余个编码ATG蛋白的基因,并且大多数在酵母和哺乳动物之间高度保守。在哺乳动物细胞中,饥饿诱导的细胞自噬大约受20种核心ATG基因调节,它们在液泡附近被不断募集,并组装形成细胞自噬前体。这些基因的分类和作用(表1)如下:
①ULK1激酶核心复合物 包括ULK1/2、ATG13、RB1CC1/FIP200和 ATG101;
②PI3K复合物 包括VPS34、VPS15、Beclin1和ATG14L;
③ATG9A转运系统 包括ATG9A、WIPI1/2和ATG2A;
④ATG12泛素样结合系统 包括ATG12、ATG7、ATG10、ATG5和ATG16L1;
⑤LC3泛素样结合系统 包括LC3A/B/C、ATG7、ATG3和ATG4A/B/C/D。
|
|
表1. 细胞自噬相关基因及其在细胞自噬中的蛋白质功能
(Li et al. Molecular Cancer,2020)
4.细胞自噬的调控
细胞基础水平的细胞自噬活性很低,不适于观察,因此,对细胞自噬研究多需人工干预。
雷帕霉素靶蛋白(mTOR)激酶在细胞自噬反应中起着重要的调节作用。mTOR(Akt and MAPK signaling)在被mTOR激酶激活后,细胞自噬反应被抑制;而在mTOR(AMPK and p53 signaling)被抑制后,细胞自噬反应机制启动。细胞自噬相关基因(autophagy related gene,Atg)通过形成Atg12-Atg5 和 LC3-II (Atg8-II)复合物来调控细胞自噬体的形成。
细胞自噬既能抑制也能促进细胞凋亡,两种反应在生物体内广泛存在。在营养缺乏时,细胞自噬反应作为一个细胞的促活反应机制存在,但是过量的细胞自噬反应也会导致细胞死亡,但由细胞自噬导致的细胞死亡与细胞凋亡在形态学上有明显的不同。几种促凋亡信号因子,如TNF、TRAIL和 FADD也会诱导细胞自噬的反应发生。此外,Bcl-2也是细胞自噬反应重要的调节因子,其为凋亡抑制蛋白,通过与Beclin-1结合形成复合物,来抑制由Beclin-1诱导的细胞自噬。
已报道的部分干预药物详见表2:
| 细胞自噬激动剂 | 细胞自噬拮抗剂 |
| Rapamycin:雷帕霉素,mTOR抑制剂(最常用); | Chloroquine:氯奎宁,溶酶体抑制剂; |
| EBSS:Earle's平衡盐溶液,制造饥饿; | Bafilomycin A1:巴佛洛霉素A1,质子泵抑制剂; |
| Lithium Chloride:氯化锂,IMPase 抑制剂 | 3-Methyladenine:3-甲基腺嘌呤hVps34抑制剂; |
| Xestospongin B/C:光溜海绵素 B/C,IP3R阻滞剂等。 | NH4Cl:氯化铵,溶酶体抑制剂等。 |
表2. 细胞自噬激动剂和拮抗剂
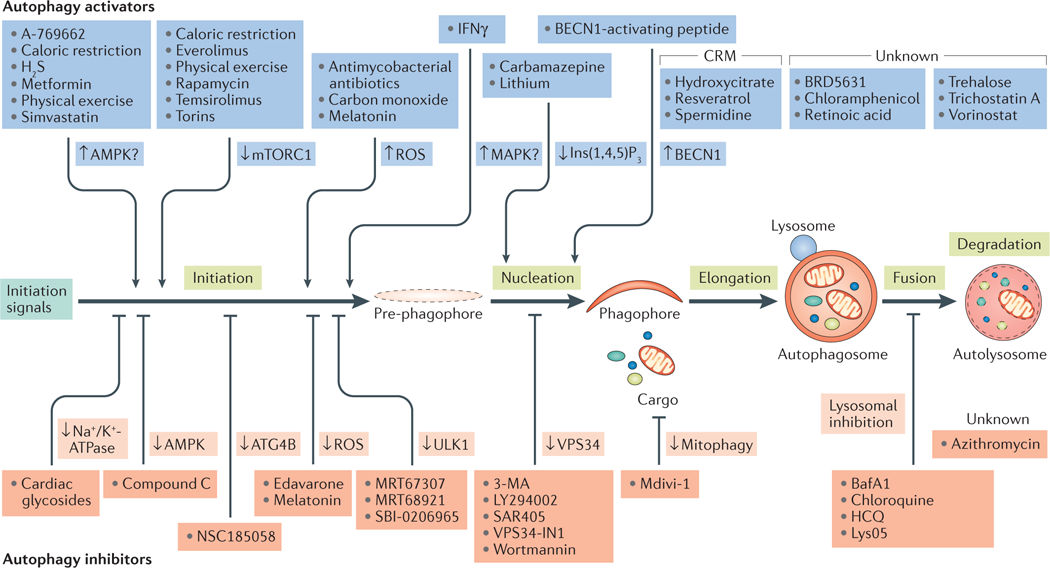
目前常用的细胞自噬激动剂有Rapamycin,EBSS等,常用的细胞自噬抑制剂有Chloroquine,3-MA,NH4Cl和 Bafilomycin A1等。这些细胞自噬激动剂和抑制剂可以分别激活/抑制细胞自噬发生的不同阶段,因此研究者可以根据实验需要选择合适的激动剂或抑制剂。更多细胞自噬通路调控药物见图3。
|
|
图3.细胞自噬通路调控药物
(Lorenzo Galluzi et al. Nat Rev Drug Discov, 2017 )
5.细胞自噬检测方法
在生理条件下,细胞自噬活性通常较低。但在饥饿、缺氧和疾病等刺激下细胞自噬活性会显著上调。另外,细胞自噬抑制也与某些疾病相关,如癌症、神经退行性疾病和传染病等。鉴于细胞自噬与不同的生理和病理生理过程之间存在紧密联系,故科学工作者需要选择理想的细胞自噬检测方法。细胞经细胞自噬诱导或抑制后,常用的观察和检测方法有∶
5.1 透射电镜法
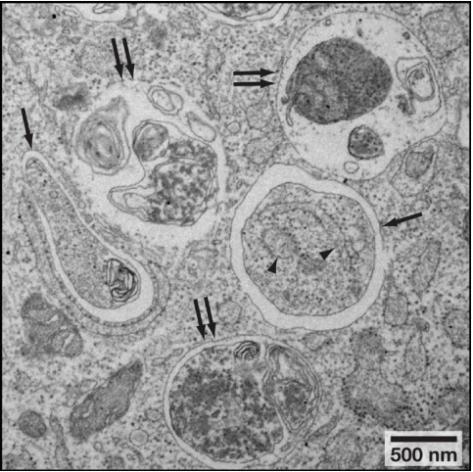
细胞自噬体属于亚细胞结构,普通光镜下看不到,直接在透射电镜下观察细胞自噬不同阶段的形态变化是一种非常直接的方法。细胞自噬各阶段的形态学特征见表3;透射电镜下细胞自噬小体和细胞自噬溶酶体的形态见图4。
| 细胞自噬阶段 | 形态特征 |
| 隔离膜 | 新月状或杯状,双层或多层膜,有包绕胞浆成分的趋势。 |
| 细胞自噬小体 | 双层或多层膜的液泡状结构,内含胞浆成分,如线粒体、内质网、核糖体等。 |
| 细胞自噬溶酶体 | 单层膜,胞浆成分已降解。 |
表3. 细胞自噬各阶段特征
|
|
图4. 透射电镜下细胞自噬小体(单箭头)和细胞自噬溶酶体(双箭头)形态
(Noboru, Mizushima et al. Cell, 2010)
5.2 荧光显微镜观察法
LC3(light chain 3)全称MAP1LC3 (microtubule-associated proteins light chain 3),贯穿整个细胞自噬过程,是目前公认的细胞自噬标记物。哺乳动物中的LC3与酵母中的细胞自噬相关蛋白Apg8/Aut7/Atg8具有同源性。哺乳动物的LC3可分为三种:LC3A、LC3B和LC3C。其中,LC3B应用最为广泛。
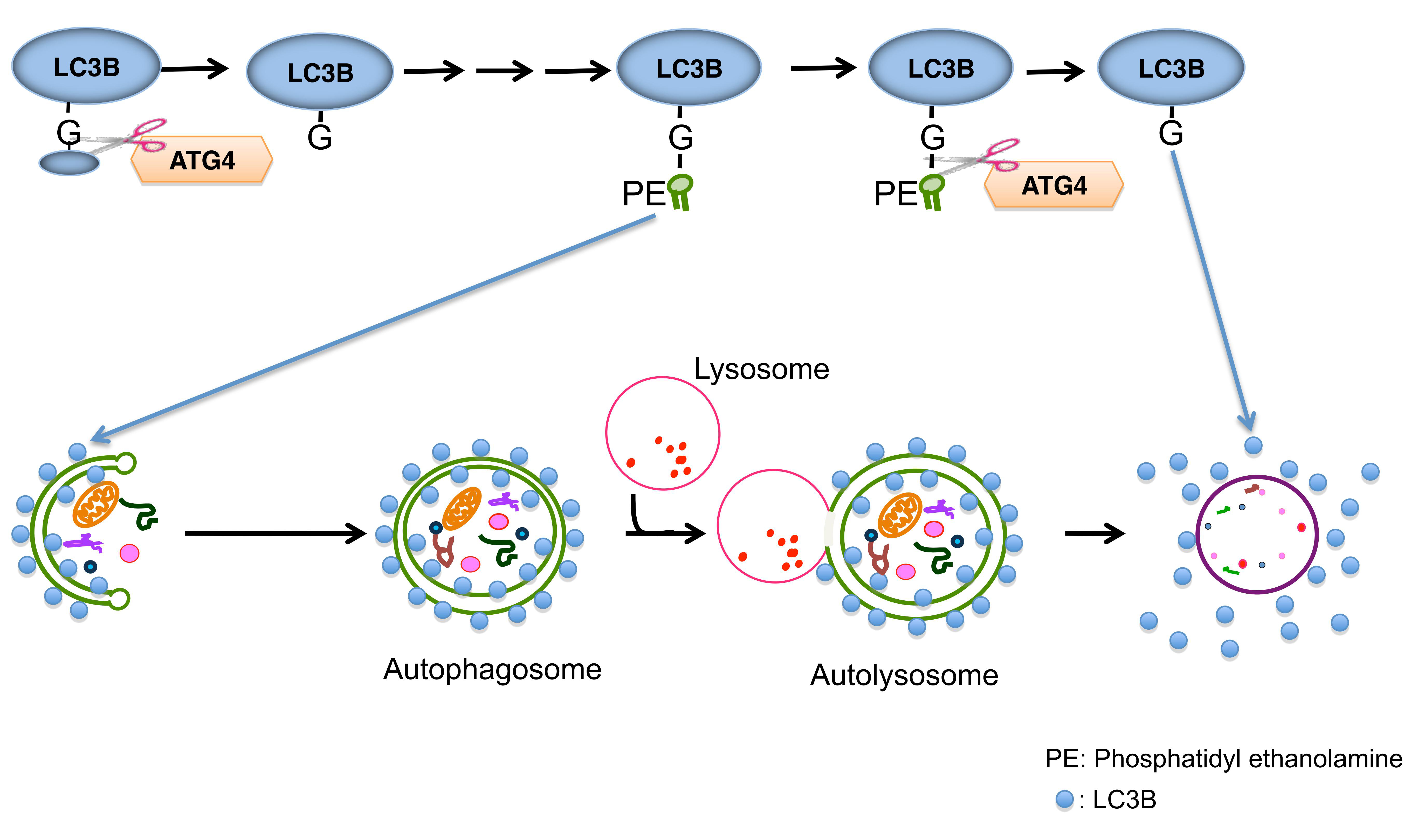
LC3蛋白合成后在其羧基端被Atg4剪切掉C端5肽,暴露甘氨酸残基,产生细胞浆定位的LC3-I。在细胞自噬过程中,LC3-I会被包括Atg7和Atg3在内的泛素样体系修饰和加工,与磷脂酰乙醇胺(PE)相偶联,形成LC3-II并定位于细胞自噬体内外膜上。细胞自噬体和溶酶体融合后,外膜上的LC3-II被Atg4切割,产生LC3-I循环利用;内膜上的LC3-II被溶酶体酶降解,导致细胞自噬溶酶体中LC3含量很低(图5)。因此,科研工作者可以通过荧光显微镜观察内源性LC3或GFP-LC3,即可实现对细胞自噬发生的检测。
|
|
图5. LC3在细胞自噬发生中的路径
(Kiriyama, Y et al. International Electronic Conference on Medicinal Chemistry,2016)
GFP-LC3单荧光和mCherry-GFP-LC3双荧光指示系统
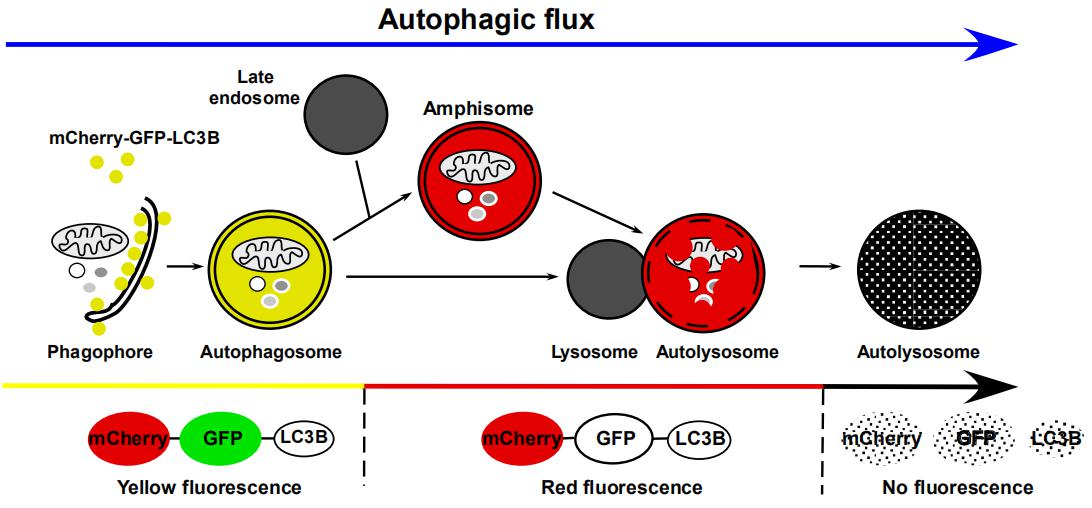
细胞自噬形成时,GFP-LC3或mCherry-GFP-LC3融合蛋白转移至细胞自噬体膜,在荧光显微镜下形成多个明亮的绿色或黄色荧光斑点。当细胞自噬溶酶体形成后,酸性的溶酶体环境使GFP荧光淬灭,而mCherry荧光不受影响,细胞自噬溶酶体呈现红色荧光(图6)。因此,科研工作者可以通过LC3荧光指示系统来监测细胞自噬流。
|
|
图6. LC3细胞自噬双标系统追踪细胞自噬流不同阶段
(Hansen T E, Johansen T. BMC Biology, 2011)
5.3 Western Blot检测LC3和p62蛋白的表达量
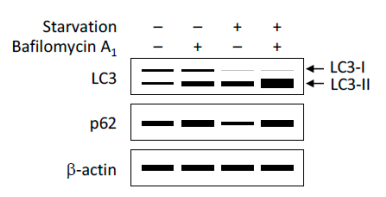
① 利用Western Blot检测LC3-II/I比值的变化以评价细胞自噬形成(图7)。细胞自噬形成时,胞浆型LC3-I会酶解掉一小段多肽,随后跟PE结合转变为膜型的LC3-II。因此可以通过LC3-II/I比值的大小估计细胞自噬水平的高低。
|
|
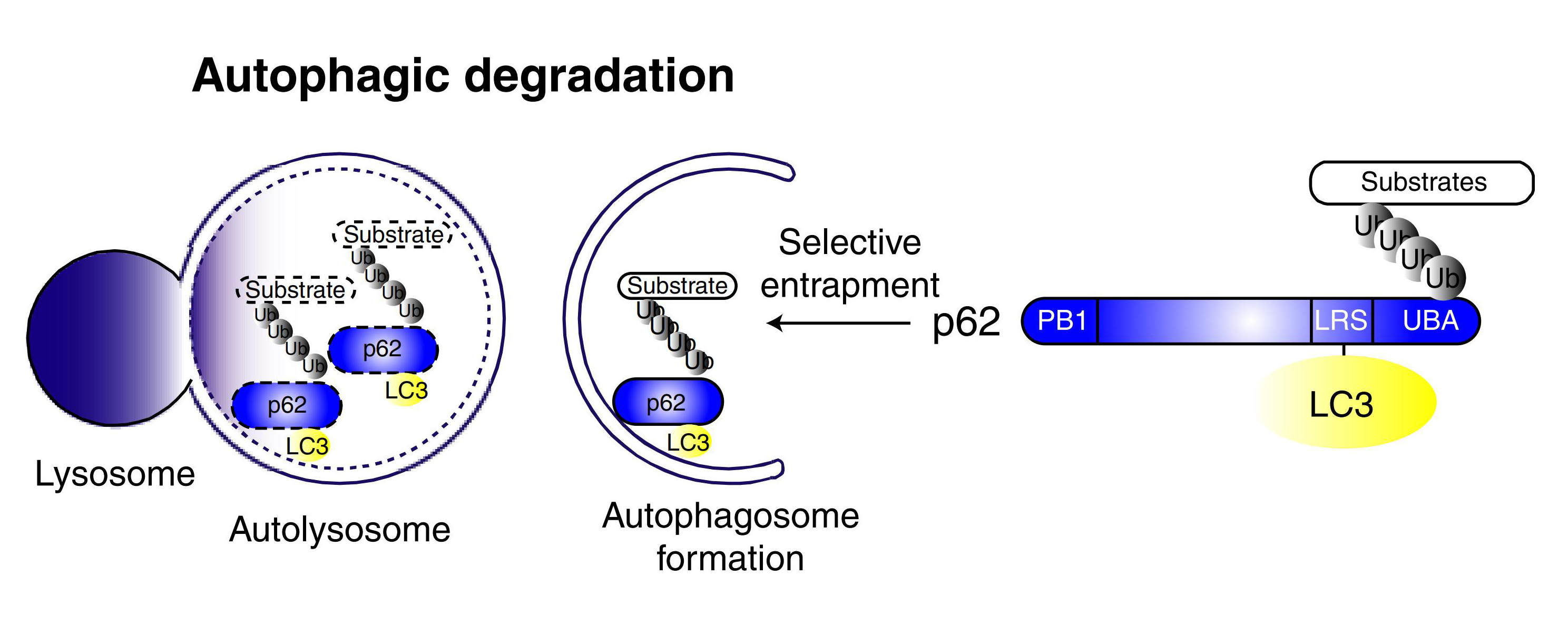
② 除LC3外,其他细胞自噬底物表达量的变化也可以用于监测细胞自噬流。其中,p62是研究广泛的一个细胞自噬底物。p62(也称为SQSTM1蛋白),由以下三个结构域组成:N端Phox和Bem1 (PB1)结构域、锌指结构域和C端泛素相关(UBA)结构域。研究表明,p62蛋白锌指结构域和UBA结构域之间的连接区域(LRS区域)负责与细胞自噬受体蛋白 Atg8/LC3结合,UBA结构域负责招募泛素化蛋白。在细胞自噬体形成过程中,p62作为链接LC3和聚泛素化蛋白之间的桥梁,被选择性地包裹进细胞自噬体,之后被细胞自噬溶酶体中的蛋白水解酶将其降解(图8),所以p62蛋白的表达量与细胞自噬活性呈现负相关。因此,利用Western Blot检测p62蛋白的表达量也可以用来评价细胞自噬水平(图7)。
|
|
图8. p62介导的选择性细胞自噬模型
(Ichimura Y et al. J Biol Chem, 2008)
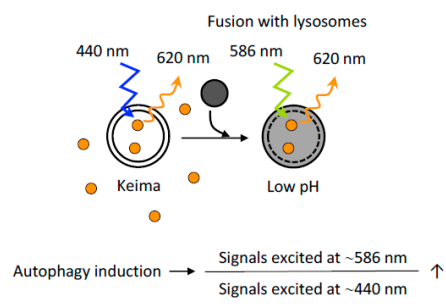
5.4 基于Keima蛋白的细胞自噬评价
Keima是一种PH敏感型的荧光蛋白,其双峰激发光谱依赖于周围的pH值,在中性和酸性环境中分别在440 nm和586 nm处激活。Keima在中性和酸性pH中荧光信号不同的特性,可以直观地反映细胞自噬程度。将细胞质的Keima传递到溶酶体可反映非选择性细胞自噬,将Keima融合到特定的蛋白(如融合到线粒体靶向序列—mt-Keima,作为线粒体细胞自噬标记)可用于反映选择性细胞自噬。值得注意是,基于keima的检测不能在固定细胞中进行,因为这种检测完全依赖于溶酶体的酸性。
|
|
6.维真细胞自噬相关产品
维真生物可提供细胞自噬研究相关工具,包含大量细胞自噬相关基因的现货克隆,shRNA克隆,CRISPR克隆,以及LC3细胞自噬单双标克隆和病毒,多种策略、多种规格、部分现货,满足您多样化的研究需求。
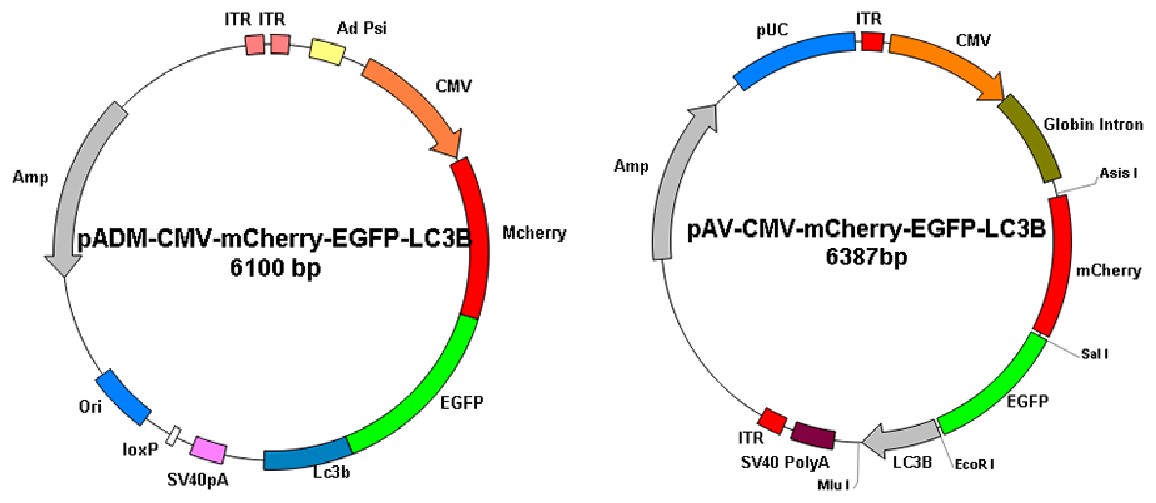
6.1 病毒产品
细胞自噬单双标病毒
| 产品名称 | 荧光蛋白 | 滴度 |
| AdV5-CMV-GFP-LC3 | GFP | ≥1*10E10pfu/ml |
| AdV5-CMV-mCherry-GFP-LC3 | mCherry、GFP | ≥1*10E10pfu/ml |
| AAV-CMV-GFP-LC3 | GFP | ≥1*10E13vg/ml |
| AAV-CMV-mCherry-GFP-LC3 | mCherry、GFP | ≥1*10E13vg/ml |
注:启动子为CMV
|
|
图10. 维真细胞自噬双标病毒载体图谱
6.2 克隆产品
部分细胞自噬相关基因
|
产品货号 |
基因名称 |
基因编号 |
基因长度 |
分类 |
|
CH842100 |
ATG13 |
NM_014741 |
1443 bp |
The ULK1 kinase core complex |
|
CH870272 |
ATG101 |
NM_021934 |
657bp |
The ULK1 kinase core complex |
|
CH826495 |
BECN1/Beclin1 |
NM_003766 |
1353bp |
The class III PI3K complex I |
|
CH810399 |
PIK3C3/VPS34 |
NM_002647 |
2664bp |
The class III PI3K complex I |
|
CH841329 |
PIK3R4/VPS15 |
NM_014602 |
4077 bp |
The class III PI3K complex I |
|
CH838864 |
ATG14/ATG14L |
NM_014924 |
1479bp |
The class III PI3K complex I |
|
CH854934 |
WIPI1 |
NM_017983 |
1341 bp |
The ATG9A/ATG2-WIPI1/2 trafficking system |
|
CH882336 |
WIPI2 |
NM_015610 |
1365 bp |
The ATG9A/ATG2-WIPI1/2 trafficking system |
|
CH821707 |
ATG12 |
NM_004707 |
423 bp |
The ATG12-conjugation system |
|
CH805558 |
ATG10 |
NM_031482 |
663 bp |
The ATG12-conjugation system |
|
CH828790 |
ATG5 |
NM_004849 |
828 bp |
The ATG12-conjugation system |
|
CH886537 |
ATG16L1 |
NM_030803 |
1824 bp |
The ATG12-conjugation system |
|
CH824179 |
MAP1LC3A/LC3A |
NM_032514 |
366 bp |
The LC3-conjugation system |
|
CH837322 |
MAP1LC3B/LC3B |
NM_022818 |
378 bp |
The LC3-conjugation system |
|
CH893675 |
MAP1LC3C/LC3C |
NM_001004343 |
444 bp |
The LC3-conjugation system |
|
CH811236 |
ATG3 |
NM_022488 |
945 bp |
The LC3-conjugation system |
|
CH878793 |
ATG4A |
NM_052936 |
1197 bp |
The LC3-conjugation system |
|
CH896350 |
ATG4B |
NM_013325 |
1182 bp |
The LC3-conjugation system |
|
CH804672 |
ATG4C |
NM_178221/ NM_032852 |
1377 bp |
The LC3-conjugation system |
|
CH800065 |
ATG4D |
NM_032885 |
1425 bp |
The LC3-conjugation system |
|
CH871029 |
ATG7 |
NM_006395 |
2112 bp |
The LC3-conjugation system |
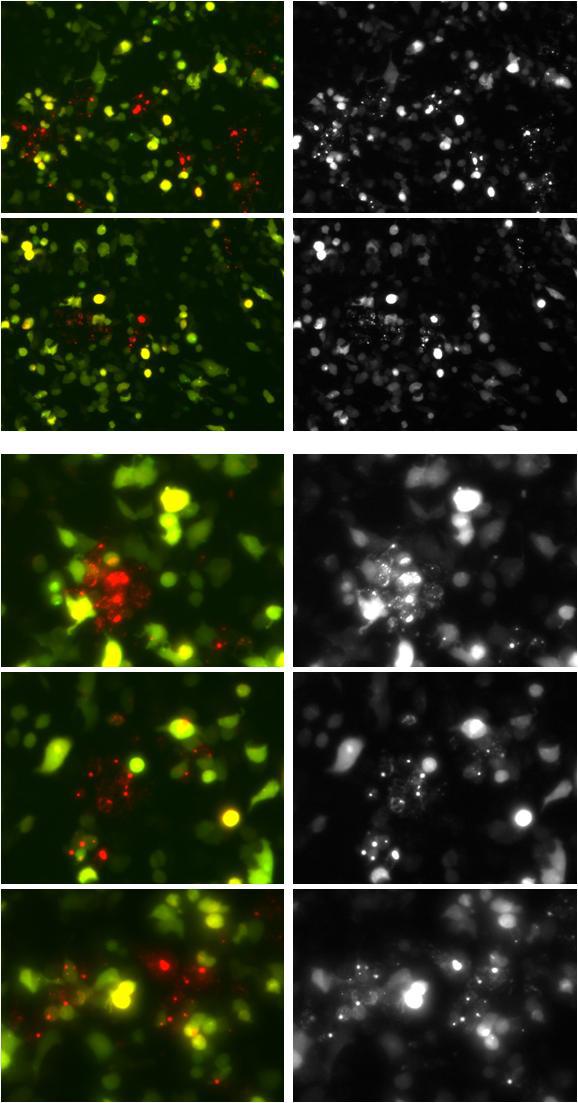
附:维真细胞自噬双标系统感染诱导细胞自噬的HEK293细胞效果图
|
|
7.客户代表性文章(部分)

400-077-2566

service@wzbio.cn